Unterabschnitte
Umbauvorgänge an der DNA
Durch Umbauvorgänge an der DNA können - wie im Fall der
Immunglobuline neue Gene entstehen oder es findet eine
Expressionsumschaltung statt, bei der ein bislang inaktives Gen
aktiviert und ein bislang aktives Gen inaktiviert wird.
Bei der Hefe findet man z.B. einen Umbau der DNA, der zu einem Wechsel
des Paarungstypus führt. Die Fähigkeit einer Hefe, sich mit einer
anderen zu Vereinigen ist von dem Paarungstyp abhängig.
Die unterschiedlichen Paarungspartner erkennen sich untereinander an
Pheromonen. Die beiden Paarungstypen werden als a und 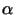 bezeichnet. Wenn sich die Zellen aneinander annähern, bleiben sie in
der G1-Phase stehen und verändern sich morphologisch. Bei einer
erfolgreichen Verschmelzung entsteht so eine diploide Zelle.
Die Verschmelzung geschieht unter der Kontrolle eines Rezeptors für
das Pheromon des jeweils anderen Typus und verläuft bei beiden Zellen
identisch in Form einer G-Protein-gekoppelten Reaktion.
bezeichnet. Wenn sich die Zellen aneinander annähern, bleiben sie in
der G1-Phase stehen und verändern sich morphologisch. Bei einer
erfolgreichen Verschmelzung entsteht so eine diploide Zelle.
Die Verschmelzung geschieht unter der Kontrolle eines Rezeptors für
das Pheromon des jeweils anderen Typus und verläuft bei beiden Zellen
identisch in Form einer G-Protein-gekoppelten Reaktion.
Diese Zelle hat die sowohl das MATa-, als auch das
MAT-Gen. Diese Zelle besitzt nun die Fähigkeit zur
Sporulation.
Bei manchen Hefestämmen hat man die Fähigkeit zeigen können, den
Paarungstyp zu wechseln. Der Besitz einest bestimmten HO-Allels macht
diesen Wechsel möglich. Dies zeigt, dass allen Zellen die Information
für beide Paarungstypen besitzen.
Dieses Phänomen wird durch ein Kassettenmodell erklärt, in dem der
MAT-Lokus eine aktive und der HML- und der HMR-Lokus eine stumme
Kassette enthält. Von diesen Genen wird nur das am MAT-Lokus
befindliche umgesetzt. Bei einem Paarungstypwechsel wird die
MAT-Kassette durch eine der stummen Kassetten ersetzt. HML enthält
nomalerweise die - und HMR die a-Kassette.
Normalerweise wird -Lokus durch ein a-Gen ersetzt und
umgekehrt.
MAT codiert für zwei Proteine 1 und 2, bzw. a1 und
a2. Diese steuern dann wiederum die Transkription der Zielgene. Dabei
werden die a-spezifischen Gene in den a Zellen konstitutiv exprimiert,
in den -Zellen jedoch reprimiert. Die -Gene werden in
den -Zellen durch Induktion exprimiert. Bestimmte
haploid-spezifische Gene werden in den haploiden Zellen exprimiert und
in den diploiden Zellen reprimiert, was dort die Expression der a- und
der -Gene verhindert.
In den MAT-Haploiden werden die a-Gene exprimiert, allerdings konnte
man ihnen in der haploiden Zelle keine Funktion zuordnen. In den
MAT-Zellen schaltet das 1-Protein die
-spezifischen Gene ein, während das 2-Protein die
a-Gene reprimiert.
In der diploiden Zelle reprimieren das a1- und das 2-Protein
gemeinsam die haploidspezifischen Gene.
An vielen Prozessen wirkt das Protein PRTF mit. Es bindet an eine
Sequenz, die P-Box und kann so die Expression der a-spezifischen Gene
aktivieren und zusammen mit 2 die a-spezifischen Proteine
reprimieren.
Bei der Expression müssen die Gene am MAT-Lokus von denen an HML und
HMR unterschieden werden. Auf beiden seiten der stummen Kassetten
liegen sogenannte Silencer, die deren Expression verhindern. Diese
verhalten sich wie negative Enhancer, d.h. sie können auch über
Distanz wirken und sie sind mit einer ARS-Sequenz verknüpft, die
normalerweise ein Replikationsorigin darstellt.
Man vermutet, dass die Silencer ihre Wirkung über eine Wechselwirkung
mit dem Chromatin entfalten. An der ARS-Sequenz bindet ein grosser
Proteinkomplex namens ORC, der normalerweise die Replikation
einleitet. Der weitere Mechanismus ist unklar.
Die selben Mechanismen, die die Transkription verhindern, verhindern
auch dass die HO-Endonuclease Zugang zu den stummen Genen bekommt und
eine Genkonversion durchführen kann.
Das HO-Gen selbst unterliegt unterschiedlichen Kontrollen: Es wird in
der diploiden Zelle nicht exprimiert; ausserdem wird es in
Tochterzellen nicht, in Mutterzellen hingegen schon exprimiert und es
wird nur am Ende der G1-Phase transkribiert. Dies bedeutet, dass ein
Paarungstypwechsel nur nach einer Zellteilung möglich ist. Nach dieser
haben die beiden Tochterzellen den zur Mutterzelle entgegengesetzten
Paarungstyp.
Trypanosomen sind die Erreger der Malaria. Dieser Erreger exprimiert
ein variables Oberflächenglykoprotein VGS, das das einzige Anitgen
seiner Oberfläche darstellt. Diese Hülle verliert der Erreger nach
seiner Aufnahme in den Darm der Fliege. dort vermehrt er sich und
seine Nachkommen haben wieder ein VSG. Diese werden dann bei dem
nächsten Stich in den Säuger abgegeben. Nachdem die Erreger dort etwa
eine Woche lang das VSG exprimiert hat, tauscht er etwa alle zwei
Wochen sein VSG. Dies wird als Antigenvariation bezeichnet.
Das VGS wird nach seiner Transkription prozessiert und mit seinem
C-Terminus in die Membran eingelagert. Dieser kann abgespalten werden,
was zur Freisetzung des Proteins führt. Die Antigenvielfalt entsteht
durch eine Expressionsumschaltung zwischen Genen. Das Basiskopiegen
verschlüsselt je eine VSG-Variante. Dabei unterscheidet man telomere
Basiskopien und interne Basiskopien. Die aktive Kopie des Gens wird
expressionsgekoppelte Kopie genannt. Diese Expressionsstelle liegt in
der Nähe eines Telomers.
Die Aktivierung eines VSG kann nun auf zwei Arten geschehen: Die
Expressionssellte kann ausgetauscht werden oder an eine andere Stelle
verlagert werden. Während die telomeren Kopien direkt aktiviert werden
könnten, wird eine interne Kopie sehr wahrscheinlich dadurch
aktiviert, dass es zunächst in eine telomere Kopie einkopiert wird und
diese dann zur Expressionsstelle wird.
Wenn der Wechsel nicht durch Aktivierung einer stummen Sequenz
geschieht, verläuft er analog der Genkonversion bei der Hefe, indem
eine stumme gegen die aktive Kassette ausgetauscht wird. Man vermutet,
dass das Aktivierungssignal eine Veränderung der Chromatinstruktur
ist.
Einige Bodenbakterien verändern das Genom einer Pflanze so, dass diese
zu einem Tumor auswächst und dem Bakterium so einen besseren
Lebensraum bietet. Diese Veränderung wird durch einen
tumorinduzierenden (Ti)-Plasmid hervorgerufen. Dieses Replicon Gene,
die für Substanzen kodieren, die von den Bakterien benötigt werden.
Das Bakterium injiziert bei der Infektion einen Teil des Ti-Plasmids,
die T-DNA in den Zellkern der Zelle - ohne dabei in die Zelle
einzudringen. Diese DNA integriert sich in das Pflanzengenom und sorgt
dort für die Transformation der Zelle und die Synthese von Opinen.
Bei dem Kontakt des Bakteriums mit der Pflanzenzelle werden dessen
vir-Gene (Virulenz-Gene) aktiviert und induzieren den Übergang der
T-DNA in die Wirtszelle. Die Übertragung der DNA in die Wirtszelle
erfolgt in Form eines Einzelstrangs und gleicht der bakteriellen
Konjugation.
In der Umgebung der Integrationsstelle findet man eine hohe Anzahl an
AT-Basenpaaren. Der genaue Integrationsprozess ist unklar. Die DNA
wird an ihrem Integrationsort exprimiert und führt zu dem
Tumorcharakter der Zelle.
Bei einer Amplifikation wird eine bereits im Genom vorhandene Sequenz
vervielfältigt und mehrfach in das Genom integriert.
Auf diese Weise entstehen häufig tandemartige Wiederholungssequenzen.
Diese Tandemanordnung kann entweder als extrachromosomale Einheit
unregelmässig vererbt werden oder stabil im Genom integriert sein. Man
kann die Genamplifikation untersuchen, indem man Zelllinen so unter
Selektionsdruck setzt, dass sie ein Resistenzgen amplifizieren. Bei
den stabilen Linien bleiben die Gene auch nach Wegfall des
Selektionsdrucks erhalten; bei den instabilen Linien verschwinden sie
danach wieder.
In den stabilen Linien lässt sich der veränderte Lokus durch eine
Anfärbung als homogen anfärbbare Region HSR sichtbar machen.
Bei den instabilen Zellen findet man keine Veränderung der
Chromosomen, sondern viele kleine double-minute-Chromosomen, die sich
autonom replizieren aber zufällig segregieren. Da diese Zellen
langsamer wachsen, verschwinden sie nach Wegfall des
Selektionsdrucks.
Die instabilen Zellen entstehen nach Einsetzen des Selektionsdrucks
zuerst. Zunächst werden dazu die chromosomalen Gene amplifiziert, dann
jedoch hängt die Resistenz und die Amplifikation der
double-minute-Chromosomen nur noch von diesen selbst ab.
Ob die instabilen Zellen durch eine verkürzte Replikation oder eine
Rekombination entstehen ist ebenso unklar wie der Mechanismus, durch
den die stabilen Zellen entstehen.
Bei einer Transfektion integriert die Zelle eine exogene DNA in ihr
Genom. Dieser Prozess entspricht der Transformation bei Bakterien.
Bei transienten oder instabilen Transfektanten integriert sich die DNA
nicht, sondern existiert als extrachromosomale Einheit. Bei einer
Integration der Sequenz in das Genom erhält man eine stabile
Zelllinie.
Die Integration der Donor-DNA scheint zufällig zu verlaufen und nicht
von einer spezifischen Sequenz abzuhängen. Häufig erfolgt eine
tandemartige Integration mehrer Plasmidkopien an eine Stelle. Wenn
ein fremdes Gen in ein Tier eingeführt wurde, wird dieses als transgen
bezeichnet. Die Aktivität des Gens ist abhängig von der
Integrationsstelle.
Normalerweise werden transgene Mäuse heute unter der Verwendung
Embryonaler Stammzellen hergestellt. Diese gewinnt man aus
Blastocysten. In diese Zelle bringt man mittels Mikroinjektion oder
Elektroporation das Gen ein. Die Zellen, in denen das Gen integriert
wurde, reichert man an und injiziert diese wiederum in eine
Blasocyste. Die Blastocyste wird in den Uterus eines Weibchens
implantiert und entwickelt sich dort weiter. Wenn man bei einer
Kreuzung der Jungtiere mit einer Maus, die das Merkmal nicht trägt,
transgene Tiere erhält, weiss man, dass man eine transgene Maus
gefunden hat, bei der das Gen stabil in die Keimbahn integriert ist.
Durch eine Weiterentwicklung diese Technik kann man gezielt Mäuse
erschaffen, bei denen durch eine homologe Rekombination das natürliche
Gen durch ein modifiziertes Homologon ersetzt wurde. Dabei verwendet
man zwei Marker, das neo-Gen (Resistenzgen) und das SV40-TK-Gen. Das
TK-Gen wird nur bei der sequenzspezifischen Rekombination, nicht bei
der homologen integriert und macht die Zelle empfindlich gegen
Ganciclovir. So kann man die Zellen, bei denen eine homologe
Rekombination stattgefunden hat, selektieren und diese dann gezielt in
eine ES einbringen.
Bei Drosophila kann man sich auch des P-Elements bedienen, um
transgene Tiere zu erzeugen. Dazu verwendet man ein intaktes und eine
defektes P-Element. Das intakte codiert für eine Transposase, die
beide Elemente erkennt und integriert. Das defekte Gen trägt dabei die
Fremd-DNA und wird im richtigen Gewebe und zur richtigen Zeit
exprimiert.
|








