Unterabschnitte
Meiose und Sexualität
Bei den höheren Eukaryonten bilden sich aus den Zellen der Keimbahn
die Gameten; Geschlechtszellen, die nur den halben (haploiden)
Chromosomensatz besitzen. Diese fusionieren dann zu einer diploiden
Zygote, aus der sich der neue Organismus entwickelt. Normalerweise ist
die diploide Phase die längste im Leben der Eukaryonten, mit der
Ausnahme von einigen Hefen, die ihr Leben normalerweise haploid
verbringen und sich direkt nach der Paarung durch eine Meiose wieder
in ein haploides Stadium begeben.
Der Vorteil der sexuellen Vermehrung liegt vor allem darin, dass es
bei einer extremen Veränderung der Umwelt eine höhere
Wahrscheinlichkeit gibt, dass sich unter den Nachkommen eine
Merkmalskombination mit einem hohen Selektionsvorteil befindet.
Als Meiose bezeichnet man die Bildung der Keimzellen bei der der
Chromosomensatz halbiert wird. Hierzu müssen die homologen Chromosomen
gleichmässig auf die zwei Zellen verteilt werden. Der Mechanismus
könnte im Prinzip wie eine Mitose ablaufen, bei der keine S-Phase
stattgefunden hat; der tatsächliche Ablauf ist allerdings wesentlich
komplexer.
Bei der Meiose kommt es nach der Replikation der DNA zu einer Paarung
der homologen Chromosomen. Die Schwesterchromatidien verhalten sich
wie eine Einheit - so als hätte die Replikation nicht statt
gefunden. Bei der Zusammenlageung der homologen Chromosomen, Bivalent
genannt, kommt es zur Rekombination zwischen dem väterlichen und dem
mütterlichen Homologon. Dann werden in einer Anaphase die
Schwesterchromatidien jeweils zusammen zu einem Spindelpol
transportiert.
So entstehen zwei diploide Zellen, die allerdings jeweils ein
verdoppeltes haploides und kein ,,echtes`` diploides Erbgut
enthalten.
Diese Zellen trennen sich nun in einer zweiten Zellteilung ohne
vorangegangene Replikation auf zwei haploide Zellen auf.
Durch das unabhängige Aufteilen der einzelnen Chromosomenpaare auf die
Zellen ergeben sich bereits 2n Möglichkeiten, wobei n die Anzahl
des halben Chromosomensatzes ist, bei uns gibt es also
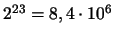 Möglichkeiten, die durch das Crossing-over nochmals
gesteigert werden. Möglichkeiten, die durch das Crossing-over nochmals
gesteigert werden.
Am Ende der ersten Teilung der Meiose liegen die Schwesterchromatiden
sehr eng zusammen und sind über das Chiasma verbunden. In diesem
Stadium können sich Bereiche der Chromosomen überschneiden und somit
Rekombinieren.
Damit diese Rekombination stattfinden kann, müssen sich die
Chromosomen zu einem sogenannten synaptonemalen Komplex
zusammenlagern. Dessen Bildung kann man in unterschiedliche Phasen
einteilen. Im Leptotän sind die homologen Chromosomen noch vollkommen
getrennt. Im Zygotän beginnt dann die Bildung des synaptonemalen
Komplexes, der sich im Pachytän stabilisiert und im darauf folgenden
Dilplotän wieder trennt. Die genetische Rekombination findet im
Parchytän während der synaptonemale Komplex existiert statt.
Man nimmt allerdings an, dass nicht der synaptonemale Koplex, sondern
die sogenannten Rekombinationsknoten für den Chromatinaustausch
verantworlich sind. Diese Knoten sind grosse Proteinkomplexe mit einem
Durchmesser von 90 nm. Man hat zwar keinen Beweis für die Rolle der
Rekombinationsknoten, allerdings deuten viele Hinweise auf den
Zusammenhang hin: Die Anzahl und die Verteilung der
Rekombinationsknoten entspricht den Chiasmata und den
Crossing-over-Ereignissen, bei Mutationen, die das Crossing-over
verändern, verändert sich auch die Lage der Rekombinationknoten und
man kann zeigen, dass an den Rekombinationsknoten eine vermehrte
DNA-Synthese stattfindet.
Ausserdem nehmen die Chiasmata die Rolle der Centromere bei der
normalen Mitose wahr und sind für die ordnungsgemässe Trennung sehr
wichtig. Während der ersten Teilung sind die Kinetochore der beiden
Schwesterchromatiden miteinander verschmolzen und die daran
angreifenden MT verlaufen alle zu einem Pol. Ausserdem bricht in der
Anaphase der ersten Meiose-Teilung nicht die Verbindung der
Schwesterchromatiden, sondern die Verbindung zwischen den homologen
Chromosomen zusammen.
Die Paarung der Geschlechtschromosomen X und Y wird durch einen
kleinen homologen Abschnitt in der Mitte dieser Chromosomen
erreicht. So können auch diese korrekt auf die Zellen verteilt
werden.
Die zweite Teilung gleicht der Mitose.
Die Eizelle
Die Eizelle eines höhern Organismus ist totpotent, d.h. sie kann einen
kopletten neuen Organismus bilden. Normalerweise geschieht dies nach
der Befruchtung, doch einige Organismen entwickeln sich auch aus
unbefuchteten Eiern (Parthenogenese, z.B. bei Eidechsen) und bei
anderen (Fröschen) kann man
diese Entwicklung durch das Einstechen der Eizelle mit einer Nadel
induzieren.
Mit Ausnahme der Säuger, bei denen das neue Individuum die Nahrung von
der Mutter erhält ist die Eizelle bei allen anderen Tiere auch
gleichzeitig eine Nahrungsreserve in Form von Dotter. Der Dotteranteil
der meisten Eier macht bis zu 95% des Volumens aus.
Praktisch alle nicht-Säugereier sind von eine speziellen Schicht, der
Eihülle umgeben, einer spezialisierten extrazellulären Matrix, die das
Ei vor mechanischen Schäden bewahrt und bei vielen Arten dient sie
auch dazu, nur die artspezifischen Spermien in die Zelle zu lassen.
Die meisten Eier enthalten sogenannte Rindengranula, die wenn das Ei
durch ein Spermium aktiviert wurde, ausgeschüttet werden und das
Eindringen weiterer Spermien verhindern.
Damit aus dem sich entwickelnden Ei, der Oozyte ein reifes Ei, ein
Ovum wird, bedarf es einer Entwicklung, die in Phasen abläuft
(Oogenese):
Die Urkeimzelle wandern zu den sich bildenden Gonaden und werden zu
Oogonien. Dann folgt eine mitotische Teilung, die aus den Oogonien die
primären Oozyten macht.
Dann beginnt die Meiose und die Zelle wird über einen Zeitraum von
einigen Tagen bis hin zu Jahren in der Prophase I der ersten Teilung
gehalten. In dieser Zeit bildet das Ei die Rindengranula, die Hülle
und es fängt bei Nicht-Säugern mit der Bildung des Dotters an (dazu
müssen die Chromosomen dekondensieren). Es steht für die Biosynthese
ein doppelter Chromosomensatz zur Verfügung, was diese sehr
beschleunigt. Die Biosynthese wird ausserdem durch die mit Gap
Junctions mit der Eizelle verbundenen Folikelzelle und teilweise durch
spezielle Nährzellen unterstützt.
Die nächste Phase nennt man die Oozyten-Reifung. Diese Phase wird
normalerweise erst nach der Geschlechtsreife durch Hormone
eingeleitet. Nun wird die erste Meioseteilung fortgesetzt, die
Chromosomen kondensieren wieder und die Kernhülle verschwindet.
Am Ende teilt sich die Zelle in zwei Zellen von sehr unterschiedlicher
Grösse: das kleine Polkörperchen und die sekundäre Oozyte.
Durch die zweite Teilung der Meiose entsteht dann aus der sekundären
Oozyte ein zweites Polkörperchen und das reife Ei (Ovum).
Bei den meisten Organismen stoppt die Reifung in der Metaphase der
zweiten Meiose-Teilung und wartet auf die Befruchtung. Diese werden
dann durch die Ovulation freigesetzt und erwarten die Befruchtung.
Die Spermienzelle
Die Spermienzelle ist eine Zelle, die an die Bewegung angepasst ist.
Sie besitzt eine Schwanzregion, die das Spermium antreibt und ihm
hilft, die Eihülle zu durchbohren, sowie einem Kopf, der das dicht
gepackte Genom enthält, bei dem aber der gesamte Apparat der
Proteinbiosynthese fehlt.
Am Vorderende des Spermiums findet man spezielle Vesikel mit
hydrolysierenden Enzymen, die die Eihülle auflösen: die
Akrosomen-Vesikel. Diese werden durch Kontakt mit der Eizelle frei und
ermöglichen dem Spermium so, die äussere Eihülle zu durchdringen.
Die Geissel des Spermiums enhält in ihrem Kern die typische
9+2-Anordnung, die jedoch von neun weiteren dichten Keratinfasern
umgeben sind, deren Rolle nicht klar ist.
Im Gegensatz zu den Oogonien der Frauen, die nur im Fötus gebildet
werden und dann in periodischen Abständen abgegeben werden, bildet der
Mann erst ab der Pubertät - dann aber kontinuierlich - Spermien.
Die Spermatogenese findet in den Samenkanälchen statt.
An deren Peripherie befinden sich die unreifen Spermienzellen - die
Spermatogonien. Einige dieser Zellen differenzieren sich zu primären
Spermatozyten. Diese beginnen dann die Meiose und teilen sich in zwei
sekundäre Spermatozyten mit jeweils einem doppelten Chromosomensatz.
Durch die zweite Mitotische Teilung entstehen vier Spermatide mit
einem haploiden Chromosomensatz.
Während der gesamten Spermatogenese bleiben die Zellen durch
cytoplasmatische Brücken zu einem Syncytium verbunden, bis dann die
fertigen Spermien synchron freigegeben werden.
Dieser Mechanis führt dazu, dass der gesamte Prozess der Spermatogenes
von einem diploiden Chromosomensatz gesteuert werden kann.
Die Befruchtung
Wenn es nicht innerhalb von einigen Stunden zur Vereinigung von
Spermium und Eizelle kommt, dann sterben beide Zellen unweigerlich
ab.
Damit es zur Befruchtung kommt muss das Spermium zunächst den Saum aus
Follikelzellen durchwandern, dann an die Eihülle, die Zona pellucida
binden und schliesslich mit der Plasmamembran verschmelzen.
Damit das Spermium nach der Ejakulation zu diesen Leistungen fähig
ist, muss es den von weiblichen Sekrete eingeleiteten Prozess der
Kapazitation durchmachen. Bei diesem Prozess werden die Glykoproteine
und die Plasmamembran des Spermiums verändert und dessen Beweglichkeit
erhöht sich.
Nachdem das Spermium die Follikelzellen durchdrungen hat bindet es an
die Zona pellucida. Diese ist normalerweise eine Befruchtungsbarriere
zwischen verschiedenen Arten.
Die Zona pellucida der Säuger ist aus drei Glykoproteinen
aufgebaut. Zwei davon, Z2 und Z3 bilden Filamente, die von Z1 zu einem
Netz verknüpft werden. Z3 ist ein artspezifischer Rezeptor, an den das
Spermium bindet. Z3 startet ausserdem die Akrosomenreaktion.
Der Inhalt des Akrosoms wird freigesetzt und dies führt zum einen zur
Freisetzung von Enzymen, die die Zona pellucida abbauen und zum
anderen werden dadurch Proteine an die Spermienoberfläche gebracht,
die dieses an den Z2-Molekülen der Zona pellucida fest verankern.
Nach dem Verschmelzen der Plasmamembranen kommt es zu einer
Depolarisation der Eizelle was weitere Spermien von einem Eindringen
abhält. Neben diesem primären Block der Polyspermie gibt es ausserdem
einen sekundären Block, der als Rinden-Reaktion bezeichnet wird. Diese
IP3-abhängige Reaktion setzt die Rindengranula frei und führt zu einer
Verhärtung der Zona pellucida, was ein weiteres Eindrigen von Spermien
unmöglich macht. Wenn doch einmal zwei Spermien eingedrungen sind,
bilden sich mehrere Spindelapparate aus und die weitere Entwicklung
der Zelle wird gestoppt.
Bei der Fusion der beiden Zellen stülpen sich die Mikrovilli der
Eizelle nach aussen und umschliessen das Spermium bis es schliesslich
vollständig mit der Zelle verschmilzt. Die Fusion wird sehr
wahrscheinlich durch die Wechselwirkung zwischen einem
Oberflächenprotein des Spermiums und einem Integrin der Plasmamembran
vermittelt.
Damit das Befruchtete Ei - Zygote genannt - entsteht müssen sich
noch die Kerne der beiden Zellen vereinen. Das Spermium enthält in den
meisten Tieren neben der DNA auch ein Centriol, welches es in die
Zelle einbringt; denn die Eizelle besitzt zwar ein Centrosom,
allerdings keine Centriolen. Nach der Verdoppelung bildet dieses die
erste mitotische Spindel aus und die Entwicklung beginnt.
|








