Unterabschnitte
Visuelles System
Photorezeptoren
Unser Wahrnehmungsbereich ist im unteren Bereich auf 300 nm
beschränkt. Die beruht darauf, dass die Athmosphäre und die Luft
bereits einen grossen Teil der kurzwelligen Strahlen filtern und
grosse Augen das kurzwellige Licht weiter streuen, so dass eine
UV-Wahrnehmung nur bei sehr kleinen Augen möglich ist. Ausserdem
bringt das UV-Licht den Glaskörper und die Linse zur Floureszenz, was
die Wahrnehmung zusätzlich behindern würde.
Im Langewellenbereich ist unsere Wahrnehmung auf 700 nm begrenzt. Die
beruht auf der Gleichung
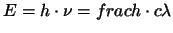 . Die Energie der Strahlung nimmmt mit zunehmender
Wellenlänge ab, so dass ein ein System, das in diesem Bereich
wahrnimmt auch durch thermisches Rauschen erregt würde. Des weiteren
würde die Augenmedien das Licht absorbieren, die Winkelauflösung
würde abnehmen und da die Rezeptoren grösser sein müssten, wäre nur
eine geringere Auflösung möglich. . Die Energie der Strahlung nimmmt mit zunehmender
Wellenlänge ab, so dass ein ein System, das in diesem Bereich
wahrnimmt auch durch thermisches Rauschen erregt würde. Des weiteren
würde die Augenmedien das Licht absorbieren, die Winkelauflösung
würde abnehmen und da die Rezeptoren grösser sein müssten, wäre nur
eine geringere Auflösung möglich.
Generell ermöglichen die Linsenaugen ein besseres Formensehen und eine
höhere Auflösung (ein Komplexauge mit der Winkelauflösung des
Linsenauges des Menschen hätte einen Druchmesser von 1 m), während
Komplexaugen ein besseres Bewegungssehen vermitteln.
Die Linsenaugen sind durch eine Linse mit einer einer Brennweite f
(die Einheit Dioptrien durch 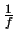 in M-1 definiert). in M-1 definiert).
Die Winkelauflösung eines Linsenauges ist duch die Beugungsphänomene
(bedingt durch die Welleneigenschaft des Lcihts) begrenzt.
In der Retina befindet sich die Foeva (siehe 5.7.2). Deren
Auflösung ist der Qualität der Linse angepasst. Die Linse bildet auch
nur in diesem Bereich ein scharfes Bild ab; da jedoch nach aussen hin
die rezeptiven Felder grösser werden, ist die irrelevant. Im Dunkeln
werden die sphärischen Fehler der Linse grösser, allerdings werden
dann auch die rezeptiven Felder der Ganglienzellen grösser, so dass
der Effekt kompensiert wird. Auch die chromatische Abberation für
blaues Licht macht sich nicht bemerkbar, da in der Fovea die
Blaurezeptoren fehlen.
Das Farbsehen beim Menschen hat neben der Farbe den Vorteil, dass es
Schärfer ist, da sich in der Fovea keine Konvergens findet und es hat
eine bessere zeitliche Auflösung.
Die Stäbchen hingegen sind empfindlicher, achromatisch, haben eine
geringe Auflösung, dafür allerdings eine esentlich höhere
Empfindlichkeit, bei der ein Photon bereits ausreicht, um eine
Reaktino auszulösen. Mehrer Stäbchen konvergieren auf eine
Bipolarzelle.
Bei den Komplexaugen fällt das Licht von einem Punkt auf je einen
Rhabdomer in sieben Ommatidien. Sechs dieser Axone konvergieren im
ersten optischen Ganglion, während das siebte Rhabdomer getrennt
weiter verarbeitet wird.
Retina
Die Vertebrantenretina besteht aus Sinnes-, Bipolar-, Ganglien,
Horizontal- und amakrinen Zellen. Das Rezeptorpotential der
Sinneszellen wird auf Neurone zweiter Ordnung übertragen.
Unter den Ganglienzellen der Retina gibt es ON-Zellen, die Durch Licht
im Zentrum erregt werden und solche, die durch Licht im Zentrum
gehemmt werden. So können Hell- und Dunkelreize übermittelt werden
ohne dass eine Zelle daueraktiv sein muss (siehe hierzu aber auch
7.5.1).
In der der Mitte der Retina befindet sich die Sehgrube, die Fovea ode
Foveola. Hier sind die Nerven zur Seite verlagert und ermöglichen so
ein besonders schafes Sehen it hoher Auflösung. Die Augen- und
kopfbewegungen sind immer darauf ausgerichtet, das interessante Objekt
auf der Foveola abzubilden.
Der bilde Fleck (Papille) markiert den Punkt, an dem die Neven in das
Auge eintreten.
Phototransduktion
Die Phototransduktion kann bereits durch die Absorption eines
einzelnen Lichtquants gestartet werden. Durch das Licht findet an
einem Chromatophor (einem Retinalderivat) eine Isomerisierung von
11-cis nach all-trans statt.
Durch diese Änderung erfährt auch das an den Chromatophor gekoppelte
Opsin (der Komplex heisst Rhodopsin) eine Konformationsänderung und
interagiert mit
Transducin. Dieses ist G-Protein-gekoppelt und löst weitere
Enzymkaskaden aus.
Bei den Vetebraten hat es den Effekt, dass der cGMP-Spiegel durch die
Hydrolyse einer Photodiesterase (ein Rhodopsin kann 105 Moleküle
ccGMP zur Spaltung bringen) sinkt,
dieses von den Kanälen abdiffundiert und sich die Natrium- und
Calciumkanäle schliessen. Dies führt zu einer Hyperpolarisation dr
Zelle. Das Transducin deaktiviert sich durch GTP-Hydrolyse.
Die Reisomieriseung bei Vertebraten geschieht, indem der Chromatophor
vom Opsin abgespalten wird und in das Pigmentepithel transportier
wird, wo er dann Reisomerisiert wird.
Bei den Invertebraten bindet ein Transmitter an den Kanal, welcher
sich darauf für Natrium und Calcium öffnet und zu einer Depolarisation
führt.
Bei den Invertebraten wird das sogenannte Metarhodopsin durch die
Aufnahme eines Lichtquants photo-reisomerisiert.
Alle vier Sehfarbstoffe enthalten Retinal in gleicher Form,
unterscheiden sich jedoch durch das Opsin.
Eine Farbwahrnehmung ist nur durch die neuronale Verrechnung mehrer
Rezeptortypen (di-, tri- oder tetrachromatisches Sehen) möglich.
Bei eirn Farbblindheit (rezessiv X-chromosomal) wird zumeist einer
oder mehrere der drei Zapfenfarbstoffe nicht gebildet.
Um im Gehirn eine Helligkeits und Farbkonstanz zu berechnen benötigt
man mehrere Rezeptoren.
Photoadaptation
Rezeptoren können dunkel- oder helladaptiert sein. Durch eine
veränderte Calciumkonzentration steigt die Aktivität der
Guanylatzyklase an; so erhöht sich die cGMP-Konzentration und die
Kanäle Öffnen sich.
Sehr helles Licht hyperpolarisiert die Zelle auf ca. -70 mV, das
Gleichgewichtspotential von Kalium. Bei Anhaltender Beleuchtung
erfolgt eine Depolarisation auf -40 mV. Mit dr Zeit ändert sich dann
die Calciumkonzentration so, dass eine Adaptation erreicht wird.
Des weiteren kann sich die Pupille schliessen und bei Fischen gibt es
die Möglichkeit, dass die Sinneszellen weiter im Epithel ,,
versenkt`` werden.
Beim Komplexauge verlagern sich die Pigmentzellen.
Bei der Adaptation des menschlichen Auges kommt es zu dem sogenannten
Kohlrausch Knick, der dann auftritt, wenn das sehen in der
Dämmerung (phtotopisches Sehen) für das die Zapfen verantwortlich sind,
in das skotopische Sehen der Stäbchen übergeht. Die Stäbchen haben
längere Aussenglieder, absorbieren deshalb mehr Licht und sind demnach
empfindlicher.
Weiterleitung des Signals
Wie schon unter 5.7.2 erwähnt, wird der Output an die
sogenannten Ganglienzellen weitergegeben. Bevor er diese erreicht, wird er
von Bipolar-, Horizontal- und Amakrinzellen verschaltet.
Die Ganglienzellen haben runde rezeptive Felder, die in zwei Bereiche unterteilt
sind: Das Zentrum und das Umfeld des rezeptiven Felds.
Werden das Umfeld und das Zentrum unterschiedlich beleuchtet, reagiert die
Ganglienzelle optima - wobei es, wie oben erwähnt, solche gibt, die auf Licht
im Zentrum und eine dunkles Umfeld und solche die auf die gegenteiligen
Lichtverhältnisse reagieren. Die Ganglienzellen reagieren aus diesem Grund
mehr auf Kontraste denn auf Intensität.
Es gibt zwei Typen von Ganglienzellen: Die M-Typen (magnus für gross), die
ein grosses rezeptives Feld haben und seltern sind und den P-Typ (parvus für
klein), die häufiger auftreten und ein kleineres rezeptives Feld besitzen.
Ausserdem gibt es sogenannte K-Zellen, die spezialisierte Felder für Kontrast,
Farbe, Richtung, etc. besitzen.
Diese Typen von Ganglienzellen unterscheiden sich auch morphologisch:
Die M-Zellen sind sogenannte Alphpzellen, die grosse Somata, weitreichende,
dicke Dendriten und dicke Axone besitzen.
Die P-Zellen haben kleine, feine Dendritenbäume; Soma und Axon sind mittelgross.
Man nennt sie Betazellen.
Die K-Zellen oder Gammazellen haben kleine Somata, dünne Axone und weniger,
aber weireichende Denriten.
Die Axon der Ganglienzellen bilden den Sehnerv.
Die Sehinformatin wird über das Corpus geniculatum laterale (CGL) an den
Thalamus, von dort an den Colliculus superior im Mesencephalon und von
dort in den Hypothalamus weiter geleitet.
Verarbeitung im ZNS
Der Sehvorgang wird normalerweise durch Sakkaden, ruckartige
Augenbewegungen eingeleitet. So richtet sich der Blick auf das Objekt.
Während der Sakkaden ist die Wahrnehmung unterdrückt, so dass man
z.B. im Spiegel immer nur dir ruhigen Augen sieht.
Durch Blickfolgebewegungen wird das Objekt auf einem Punkt der Retina
fixiert - auch wenn es sich bewegt.
Insekten führen Körpersakkaden aus, da ihre Augen fest mit dem Kopf
verbunden sind.
Dann wird das Licht über die Retina wie in 5.7.2 schrieben,
durch die Retina aufgenommen und an die Ganglienzellen weitergeleitet
(siehe 5.7.5).
Der Sehnerv projiziert nun streng retinotop ind in das CGL (Corpus
geniculatum laterale). Dieses liegt im Thalamus und ist in Magno-,
Parvo- und Koniozelluläre Schichten unterteilt. Das CGL kann durch
den Cortex moduliert werden.
Insgesamt enthält der Cortex 32 Repräsentationen der Retina.
Der visuelle Cortex (V1) ist morphologisch in sechs Schichten aufzuteilen.
In Schicht II und III befinden sich die Somata der Pyramidenzellen. Diese
Schichten sind der Ausgangpunkt für inter- und intrazeluläre Projektionen.
Die Schicht IV ist am auffälligsten; dort enden die M- und P-Fasern.
In Schicht V befinden sich grosse Pyramidenzellen, die kortikofugal projizieren.
Schicht VI ist für dioe Rückprojektion zum CGL verantwortlich.
Senkrecht zu diesen Schichten befinden sich die Augendominanzsäulen.
Färbt man die Area V1 mit Cytochromoxidase an, so zeigen sich
charakteristische ,,blobs``, Strukturen mit ca. 0,2 mm Durchmesser.
Die zwischen diesen Regionen liegenden Bereich werden als Interblob-Regionen
bezeichnet.
Die selbe Färbung bei der Area V2 produziert ein Muster aus Streifen- und
Interstreifen-Regionen
Wahrnehmung
Die visuelle Wahrnehmung verläuft prinzipiell in zwei Phasen:
In der präattentiven Phase wird eine schnelle Überprüfung der Objektgrenzen
durchgeführt, während bei der attentiven Wahrnehmung feinere Merkmale
fokussiert werden.
Die Wahrnehmung verläuft in Bahnen:
- Die Parvo-Blob-Bahn ist für die Farbwahrnehmung verantwortlich.
- Die Parvo-Interblob-Bahn regelt die Formwahrnehmung.
- Eine dritte Bahn ist für die Wahrnehmung von Bewegung und
räumlichen Beziehungen verantwortlich.
Man kann über verschiedene Versuche zeigen, wie die Informationen über die
Bahnen verteilt sind. So zeigt sich z.B. durch Versuche mit äquilumineszenten
Bildern, bei denen sich die Farben, aber nicht deren Helligkeiten unterscheiden,
dass Bewegungen nur auf Grund der Helligkeit wahrgenommen werden.
Für die Objekt- und Bewegungsdetektion werden Bilder in Objekte (nach Farbe,
Form, Textur, gemeinsamer Bewegungsrichtung) zerlegt.
Interessante Untersuchungen kann man auch bei Menschen mit Agnosien
machen. Bei diesen Krankheiten sind bestimmte Wahrnehmungsaspekte
gestört. Der Patient kann z.B. keine räumlichen Zusammenhänge mehr
wahrnehmen (Tiefenagnosie), kann keine Bewegungen mehr wahrnehmen
(Bewegungsagnosie) oder er hat keine Gesichtserkennung mehr
(Prosopagnosie).
Der Cortex ist verantwortlich für die Integration verschiedener Merkmale
wie Farbe, Form, etc. Für dieses Zusammenfügen ist Aufmerksamkeit
notwendig.
Die Neurone im Nucleus praetectalis olivaris (NPO) haben Entladungsraten,
deren Logarithmus proportional der Helligkeit ist.
Der Nucleus suprachiasmaticus (NSC) liegt über dem Chiasma und reagiert
tonisch auf Erhöhung der Helligkeit (unterdrückt) oder Erniedrigung der
Helligkeit (erhöht) und regelt so den 24-h-Rhythmus (siehe
7.8).
Das Okzipitio-parietale System verläuft von IVB zu den Streifen der V2-Region ,
von dort nach V3 und von da nach V5. Die Bewegung wird über
bewegungssensitive Neurone verfolgt.
Bei Insekten gibt es in der Lamina Säulen, die jeweils einem Ommatidium
entsprechen. Die Laminasäulen projizieren in die Medulla, wo es zu
einer lateralen Wechselwirkung kommt.
In der proximalen Medulla findet man Felder, die sensitiv für die
Geschwindigkeit und den Betrag von Reizen sind.
Von den Medullasäulengehen Neuron zur Lobulaplatte und enden dort in
Richtungsschichten.
Im Chiasma kreuzen dich die optischen Nerven der beiden Augen. Bei den
Sauropsiden werden die Bilder der Augen ausschliesslich kontralateral
abgebildet, bei den Säugern und Amphibien werden die meisten Fasern
kontrlateral abgebildet; nur enige der temporalen Fasern kreuzen nicht
und bei den Primaten und eineigen Flughunden ist es so, dass die
nasalen Fasern kotra- und die temporalen Fasern ispilateral
verarbeitet werden. So werden die von einen Punkt kommenden
Informationen gemeinsam verarbeitet.
Jeder der Zapfen reagiert mit einer anderen Wahrscheinlichkeit auf ein
Signal bestimmter Wellenlänge. Bei einem Zapfen vom R-Typ kann also
ein schwaches Rotes Licht den gleichen Effekt erzeugen, wir ein
starkes Blaues Licht.
Auf Grund des Phänomens, dass man bestimmte Farben nicht in
Kombination sehen kann (es gibt kein rötliches Grün), schloss man dass
Signale mit unterschiedlicher Spektralempfindlichkeit werden
neural antagonistisch verschaltet werden (Gegenfarbtheorie).
Diese Gegenfarben oder Farbantagonisten sind Grün und Rot, Gelb und
Blau sowie Schwarz und Weiss.
Sind beide ausbalanciert, gibt es kein Signal.
Auf die Gegenfarbtheorie geht auch der Farbsimultankontrast
zurück. Wenn die Gegenfarbe von benachbarten Stellen (d.h. den
Hintergrund) ausgeht, tritt das Objekt stärker hervor.
Durch die Berechnungen des Gehirns wird weiterhin eine Farbkonstanz
geschaffen. Dies sorgt dafür, dass wir eine Farbe auch bei
unterschiedlicher Beleuchtung immer als die gleiche Farbe wahrnehmen.
Die Farbinformation wird in das Gehirn über zwei Bahnen
weitergeleitet: Über die konzentrischen Breitbandzellen und die
Gegenfarbzellen.
Die konzentrischen Breitbandzellen enthalten keine
Farbinformationenen, sondern nur einen Helligkeitswert.
Die Gegenfarbzellen sind Ganglienzellen vom Typ M oder P und
verarbeiten die Informationen der R- und G-Zellen antagonistisch. Es
kommt zu einer besonders starken Erregung, wenn das Umfeld die
Gegenfarbe des Zentrums erhält. Diese Zellen sind einfache
konzentrische Gegenfarbzellen.
Die Informationen der B-Zellen stehen in dem gesamten Feld den
Informationen aus den beiden anderen kombinierten Werten gegenüber.
Die Informationenen fliessen in V1 zusammen und werden dort auf
doppelte, konzentrische Gegenfarbzellen verschaltet. Diese Zellen
reagieen besonders gut auf Grün vor Rot, auf Blau vor Gelb und auf
Gelb vor Blau.
Von dort folgt eine Verschaltung nach V2 und von dort nach V4.
Die Formerkennung findet im visuellen Cortex statt.Bei den beteiligten
Zellen unterscheidet man zwei Arten: einfache und komplexe Zellen.
Die einfachen Zellen konvergieren auf eine komplexe Zelle.
Die Zellen sind in sogenannten Orientierungssäulen
angeordnet. Jede dieser Säulen ist ca. 30 bis 100 m breit und
enthält zum einen 4C-Zellen mit konzentrischen rezeptiven Feldern und
zum anderen in den Schichten 2, 3, 5 und 6 komplexe Zellen.
Die Säulen werden durch blobs (s.o.) für die Farbanalyse
unterbrochen und sind horizontal miteinander verknüpft. Jeder
Orientierungssäule ist eine Richtung zugeordnet; eine Gruppe von
Orientierungssäulen, die 360 abdeckt bildet zusammen mit einer linken
und einer rechten Augendominanzsäule für das binokulare Sehen und
mehreren blobs eine Hyperkolummne mit 1 mm2 Oberfläche.
Die einfachen Zellen erhalten ihre Informationen von dern Sternzellen
in Schicht 4C und reagieren auf einfache Reize, wie
z.B. Lichtpunkte. Sie haben rechteckige rezeptive Felder, die auf
einen bestimmten Reiz reagieren. und reagieren auf einfache Reize, wie
z.B. Lichtpunkte. Sie haben rechteckige rezeptive Felder, die auf
einen bestimmten Reiz reagieren.
Die komplexen Zellen haben grössere und komplexere rezeptive Felder
und reagieren positionsinvariant (Merkmale werden unabhängig von ihrer
Ausrichtung erkannt) auf Balken und Kanten.
Die Gesichtserkennung verläuft über das Parvo-Interblob-System
(d.h. von dem CGL aus zum visuellen Cortex V1, von dort nach V2 und
von dort nach V4 und von V4 zum inferior-temporalen Cortex).
Während die Zellen in V4 noch retinotop organisiert sind, findet man
im inferior-temporalen Cortex keine retinotope Organisation. Einige
Zellen antworten allerdings selektiv auf Gesichter.
Bei der Bewegungserkennung arbeiten zwei Systeme zusammen. Das eine
basiert auf Bewegungen auf der Retina, das andere basiert auf den
Bewegungen von Kopf und Augen.
Durch das begrenzte Bewegungssehen des Linsenauges kann man
Scheinbewegungen, wie z.B. in Filmen erzeugen.
|








